LYME TRANSMISSION: Transmission and Maintenance
|
Transmission& Maintenance:
In this Section: * How Lyme is Transmitted and Maintained within the environment *Ticks and Human Disease *Table of Tick Vectors of Lyme For ease of access this table has also been reproduced as a stand alone table. Click here |
|
Lyme Disease : Transmission and Maintenance
Lyme Disease (LD) / Lyme Borreliosis is described as a vector-borne disease as it is spread via the bite of arachnids (ticks). It should be noted that there are also various other methods of transmission, some confirmed, and others requiring further research. These methods include:
1. Evidence that blood sucking insects (other than ticks) are able to transmit Lyme disease
2. Contact (urine passed) transmission has been noted in mice, and
3. Human to human transmission : sexual and mother to baby (vertical)
4. Organ and Blood Donation
These areas are examined in further detail (with references provided) on the Lyme Transmission : Various Transmission Methods page of this website.
In order to address the problems with the lack of recognition of Lyme, the focus of the information on this page is on the well known ability of the tick to spread and transmit Lyme Borreliosis.
1. Evidence that blood sucking insects (other than ticks) are able to transmit Lyme disease
2. Contact (urine passed) transmission has been noted in mice, and
3. Human to human transmission : sexual and mother to baby (vertical)
4. Organ and Blood Donation
These areas are examined in further detail (with references provided) on the Lyme Transmission : Various Transmission Methods page of this website.
In order to address the problems with the lack of recognition of Lyme, the focus of the information on this page is on the well known ability of the tick to spread and transmit Lyme Borreliosis.
How Lyme is Transmitted and Maintained in the Environment
The transmission and maintenance of the bacteria responsible for Lyme disease / Borreliosis within the environment requires the tick and host animals. The host animals may be thought of as either reservoir hosts, which are small to medium size animals that carry the spirochete infection within their blood and the larger host animal for which the adult of a particular species of tick has an affinity (30). There is some question as to whether or not larger mammals such as sheep, deer, horses and cattle simply serve to amplify the infection within the environment, by providing the tick with a host blood meal or whether they also serve as reservoir hosts of Borrelia. In general, the studies show mixed conclusions. These findings are discussed further in the ‘ Scrub Tick Haemaphysalis longicornis and associated Mammal Vector & Reservoir Hosts ’ section. With regards to the smaller/medium animals, there are over 50 mammalian and avian species that are reservoir hosts of Borrelia (31) and include various mammal species such as: mice, rats, voles; hares; rabbits; squirrels; hedgehogs; dogs: as well as numerous species of marine and land birds including puffins, blackbirds and pheasants.
The transmission and maintenance of the bacteria responsible for Lyme disease / Borreliosis within the environment requires the tick and host animals. The host animals may be thought of as either reservoir hosts, which are small to medium size animals that carry the spirochete infection within their blood and the larger host animal for which the adult of a particular species of tick has an affinity (30). There is some question as to whether or not larger mammals such as sheep, deer, horses and cattle simply serve to amplify the infection within the environment, by providing the tick with a host blood meal or whether they also serve as reservoir hosts of Borrelia. In general, the studies show mixed conclusions. These findings are discussed further in the ‘ Scrub Tick Haemaphysalis longicornis and associated Mammal Vector & Reservoir Hosts ’ section. With regards to the smaller/medium animals, there are over 50 mammalian and avian species that are reservoir hosts of Borrelia (31) and include various mammal species such as: mice, rats, voles; hares; rabbits; squirrels; hedgehogs; dogs: as well as numerous species of marine and land birds including puffins, blackbirds and pheasants.
Ticks and Human Disease
Ticks are classified as arachnids (eg: spiders, mites, scorpions), as they have eight legs, rather than six as with insects (32). There are approximately 850 species of ticks worldwide that are divided into two families, the Ixodidae (hard ticks) and Argasidae (soft ticks). Both are vectors for human disease, although in the case of Lyme disease it is the Ixodidae family that has been associated with transmission. The family of Ixodidae tick itself has approximately 650 different species, divided into 13 genera including; Ixodes, Amblyomma, Haemaphysalis, Rhipicephalus and Dermacentor (33).
The first ticks found to be competent vectors of Borrelia were of the Ixodes genera: I. scapularis (Previously known as I. dammini, before being shown to be same species), and I. pacificus (Black-legged Tick), commonly known as deer ticks in America. In Europe and Asia, the vectors were found to be the I. ricinus (Castor Bean/Sheep Tick) and I. persulcatus (Taiga Tick). Since these early investigations, many more species of ticks have been identified as vectors. This includes over a dozen more species of Ixodes ticks, as well as ticks from other Ixodidae genera’s including, Amblyomma, Haemaphysalis, Rhipicephalus and Dermacentor. (A table of these ticks is presented at the end of this section, as can be seen a number of them are in Australia).
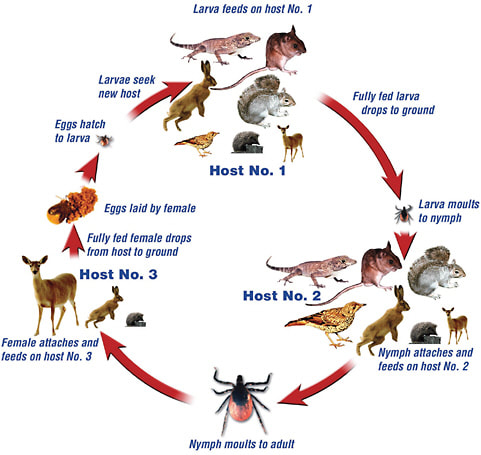
The basic implication of these findings is that there are many ticks that are capable of carrying and transmitting the bacteria that causes Lyme disease. What all of the ticks typically have in common is that they are three-host ticks. This simply means that they attach to a different host in each stage of their life development. Once the tick egg hatches to the larvae, the larvae need to find a host to attach to for a blood-meal, it then drops off and molts into a nymph. The nymph repeats the action of finding a host for a blood-meal before molting into an adult. The final blood-meal is then sought by the adult before dropping off, with the females then laying eggs. It is due to the attachment on three different hosts that these ticks are able to firstly be infected and then spread/maintain the disease within their environment.
Typically the larvae and nymphs feed on smaller animals within the environment, with the adult ticks then attaching to larger hosts (eg: deers for I. scapularis; sheep in the case of I. ricinus). It is the smaller/medium sized animals that the larvae and nymphs feed on, that act as reservoir hosts for the Borrelia bacteria that play a large role in maintaining the infectious cycle. When the larvae or nymph ticks feed on the reservoir hosts, they are then infected, and upon attaching to their next host, may pass that infection on. Whilst humans are not the preferred host, if they inadvertently come into contact with ticks, (walking through bush or long grass) then they may be at risk. It is usually at the nymphal stage that humans are infected, as at this stage of its life, the tick is barely large enough to be noticed and as ticks inject an anaesthetic into the skin of the host when attaching, the tick may feed and drop off without a person even realising.
Ticks are classified as arachnids (eg: spiders, mites, scorpions), as they have eight legs, rather than six as with insects (32). There are approximately 850 species of ticks worldwide that are divided into two families, the Ixodidae (hard ticks) and Argasidae (soft ticks). Both are vectors for human disease, although in the case of Lyme disease it is the Ixodidae family that has been associated with transmission. The family of Ixodidae tick itself has approximately 650 different species, divided into 13 genera including; Ixodes, Amblyomma, Haemaphysalis, Rhipicephalus and Dermacentor (33).
The first ticks found to be competent vectors of Borrelia were of the Ixodes genera: I. scapularis (Previously known as I. dammini, before being shown to be same species), and I. pacificus (Black-legged Tick), commonly known as deer ticks in America. In Europe and Asia, the vectors were found to be the I. ricinus (Castor Bean/Sheep Tick) and I. persulcatus (Taiga Tick). Since these early investigations, many more species of ticks have been identified as vectors. This includes over a dozen more species of Ixodes ticks, as well as ticks from other Ixodidae genera’s including, Amblyomma, Haemaphysalis, Rhipicephalus and Dermacentor. (A table of these ticks is presented at the end of this section, as can be seen a number of them are in Australia).
The basic implication of these findings is that there are many ticks that are capable of carrying and transmitting the bacteria that causes Lyme disease. What all of the ticks typically have in common is that they are three-host ticks. This simply means that they attach to a different host in each stage of their life development. Once the tick egg hatches to the larvae, the larvae need to find a host to attach to for a blood-meal, it then drops off and molts into a nymph. The nymph repeats the action of finding a host for a blood-meal before molting into an adult. The final blood-meal is then sought by the adult before dropping off, with the females then laying eggs. It is due to the attachment on three different hosts that these ticks are able to firstly be infected and then spread/maintain the disease within their environment.
Typically the larvae and nymphs feed on smaller animals within the environment, with the adult ticks then attaching to larger hosts (eg: deers for I. scapularis; sheep in the case of I. ricinus). It is the smaller/medium sized animals that the larvae and nymphs feed on, that act as reservoir hosts for the Borrelia bacteria that play a large role in maintaining the infectious cycle. When the larvae or nymph ticks feed on the reservoir hosts, they are then infected, and upon attaching to their next host, may pass that infection on. Whilst humans are not the preferred host, if they inadvertently come into contact with ticks, (walking through bush or long grass) then they may be at risk. It is usually at the nymphal stage that humans are infected, as at this stage of its life, the tick is barely large enough to be noticed and as ticks inject an anaesthetic into the skin of the host when attaching, the tick may feed and drop off without a person even realising.
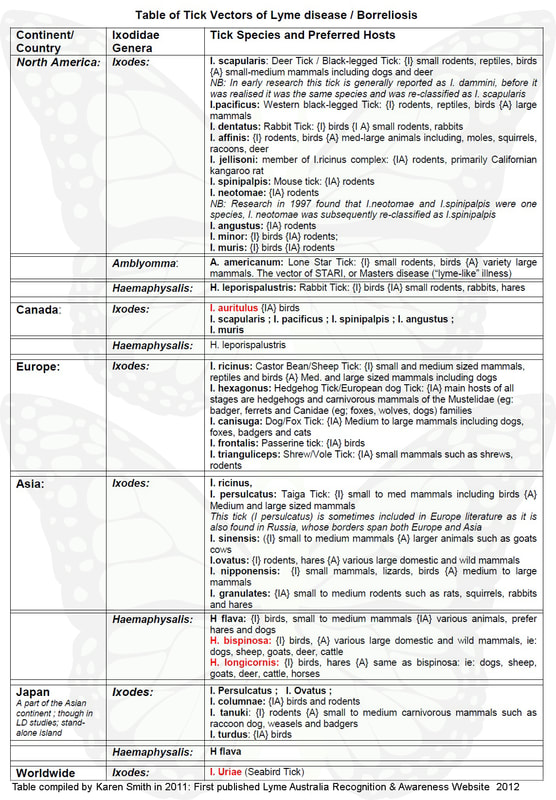
Tick Vectors of Lyme disease / Borreliosis
Below is a table of tick vectors involved in the transmission and maintenance of Lyme. The table was compiled by Karen Smith (Author of LARA) in 2011 and is by no means a fully comprehensive list of tick vectors involved in the Lyme cycle.
A key to reading the Tick Vectors of Lyme Disease/ Borreliosis Table:
The relevant ticks are listed, firstly under the country/continent in which they are found and then under their relevant Ixodidae genera, eg: Ixodes, Amblyomma, Haemaphysalis, Rhipicephalus and Dermacentor. The “scientific” name for the tick is firstly given, with the more common name (if applicable) in brackets; Animal hosts of the ticks are mentioned, with: {I} denoting hosts of Immature ticks ie: larvae and nymphs and {A} for the animal hosts of the adult ticks; If it is a second listing for the tick, that is, the tick is found in more than one continent/country, the animal hosts of the tick are not listed again. Tick species that have been recorded as being in Australia are highlighted in red.
A key to reading the Tick Vectors of Lyme Disease/ Borreliosis Table:
The relevant ticks are listed, firstly under the country/continent in which they are found and then under their relevant Ixodidae genera, eg: Ixodes, Amblyomma, Haemaphysalis, Rhipicephalus and Dermacentor. The “scientific” name for the tick is firstly given, with the more common name (if applicable) in brackets; Animal hosts of the ticks are mentioned, with: {I} denoting hosts of Immature ticks ie: larvae and nymphs and {A} for the animal hosts of the adult ticks; If it is a second listing for the tick, that is, the tick is found in more than one continent/country, the animal hosts of the tick are not listed again. Tick species that have been recorded as being in Australia are highlighted in red.
|
*See reference list below for source of tick location, animal hosts and journal articles with regards to vector capabilities of each of the above listed ticks (referenced in order of mention).
**Ticks such as I. jellisoni, I. trianguliceps and I. spinipalpis are known as nidicolous ticks (found in the burrows and nests of their hosts) and as these ticks do not actively look for hosts, their roles as vectors is associated with maintaining the Borrelia (and numerous co-infections such as Babesia microti) within the environment, rather than transmitting it to humans (1-3). However, in cases where they do come into contact with people, such as with I. spinipalpis (4), transmission to humans may occur. |
The above table, though comprehensive, is not a complete list of ticks involved in the Borrelia cycle around the world. The main aim of the table is to show how many various genera of the Ixodidae tick family are involved in the Lyme disease/ borreliosis cycle. The existence of Lyme disease in Australia was denied by Russell et al., (1994) and continues to be denied by the NSW Department of Medical entomology (of which Russell was the Director until his retirement in mid 2012), and the NSW Health Director of Communicable Diseases, Dr Jeremy McAnulty, in part due to the fact that Australia does not have any of the first four ticks (ie: I. scapularis, I. pacificus, I. ricinus, I. persulcatus) that were initially identified as vectors of Lyme in the United States and Europe. As the tick table demonstrates, there are numerous tick species that are involved in the Lyme borreliosis cycle and these species can be different in various countries. Australia has a number of tick species (highlighted in red) that are involved in the cycle, and these are discussed in further detail in the following segments:
Birds as Vector and Reservoir Hosts in Borrelia (Including examination of I. uriae (seabird) & I. auritulus (bird) ticks
Mammals as Vector and Reservoir Hosts in Borrelia (Including examination of H. longicornis & (scrub/bush) & H. bispinosa ticks
Rhipicephalus and Dermecentor Ticks
Birds as Vector and Reservoir Hosts in Borrelia (Including examination of I. uriae (seabird) & I. auritulus (bird) ticks
Mammals as Vector and Reservoir Hosts in Borrelia (Including examination of H. longicornis & (scrub/bush) & H. bispinosa ticks
Rhipicephalus and Dermecentor Ticks
For Journal article references for Borrelia Vector / Tick Vector capabilities discussed, see the Reference list below.
Tick location and animal hosts in the above table were sourced from a variety of places. The majority of information is from:
List of Australian Ticks and their hosts
Fauna of Ixodid Ticks of the World (GV Kolonin 2009) Accessed 2011-2012 [ http://www.kolonin.org/2.html ]
*Update 2017 : This resource has now been archived, some of which can be seen HERE
Other sites/information accessed include:
Ticks and Tickborne Bacterial Diseases in Humans Chart Accessed from the Journal Reading: Parola P and Raoult D (2001) Ticks and Tickborne Bacterial Diseases in Humans: An Emerging Infectious Threat: Clin Infect Dis; 32(6): 897-928
The Merck Veterinary Manual
List of Australian Ticks and their hosts
Fauna of Ixodid Ticks of the World (GV Kolonin 2009) Accessed 2011-2012 [ http://www.kolonin.org/2.html ]
*Update 2017 : This resource has now been archived, some of which can be seen HERE
Other sites/information accessed include:
Ticks and Tickborne Bacterial Diseases in Humans Chart Accessed from the Journal Reading: Parola P and Raoult D (2001) Ticks and Tickborne Bacterial Diseases in Humans: An Emerging Infectious Threat: Clin Infect Dis; 32(6): 897-928
The Merck Veterinary Manual
References: LYME TRANSMISSION: Transmission and Maintenance
Please note: Any information with regards to Lyme disease that is freely available at numerous locations on the internet has not been referenced. For specific facts/arguments, see the reference list.
NB: Reference section is separated into segments for ease of updating information
NB: Reference section is separated into segments for ease of updating information
References 1-29 can be seen / are relevant to the Lyme Transmission : Various Transmission Methods section.
How Lyme is Transmitted and Maintained in the Environment. Ticks and Human Disease
(30) Gern L and Humair PF (2002) Ecology of Borrelia burgdorferi senso lato in Europe. Lyme Borreliosis : Biology, Epidemiology and Control 6: 149-174 http://doc.rero.ch/lm.php?url=1000,43,4,20060515171152-TL/1_Gern_Lise_-_Ecology_of_Borrelia_burgdorferi_20060515.pdf
(31) Kurtenbach K, De Michelis S, Sewell HS, Etti S, Schafer SM, Hails R, Collares-Pereira M, Santos-Reis M, Hanincova K, Labuda M, Bormane A and Donaghy M (2001) Distinct Combinations of Borrelia burgdorferi Sensu Lato Genospecies Found in Individual Questing Ticks from Europe. Appl Environ Microbiol; 67(10):4926-9. http://www.ncbi.nlm.nih.gov/pubmed/11571205
(32) Vector-Borne Diseases ; e-notes: http://www.enotes.com/vector-borne-diseases-reference/vector-borne-diseases
(33) Depart Entomology, Uni of California: http://entomology.ucdavis.edu/faculty/rbkimsey/tickbio.html
Lyme Disease Tick Vectors
I. scapularis and I. pacificus: Well known vectors
I. dentatus:
(a) Anderson JF, Magnarelli LA and Stafford KC 3rd (1990) Bird-feeding ticks transstadially transmit Borrelia burgdorferi that infect Syrian hamsters. J Wildl Dis; 26 (1):1-10. http://www.ncbi.nlm.nih.gov/pubmed/2304189
(b) Indirect ref: Masuzawa T (2004) Terrestrial Distribution of the Lyme Borreliosis Agent Borrelia burgdorferi Sensu Lato in East Asia. Jpn J Infect Dis, 57(6); 229-235. http://www.ncbi.nlm.nih.gov/pubmed/15623946
I. affinis:
Maggi RG, Reichelt S, Toliver M and Engber B (2010) Borrelia species in Ixodes affinis and Ixodes scapularis ticks collected from the coastal plain of North Carolina. Ticks Tick Borne Dis;1(4):168-71. Epub 2010 Oct 20. http://www.ncbi.nlm.nih.gov/pubmed/21771524
I. jellisoni:
Lane RS, Peavey CA, Padgett KA and Hendson M (1999) Life history of Ixodes (Ixodes) jellisoni (Acari: Ixodidae) and its vector competence for Borrelia burgdorferi sensu lato. J Med Entomol;36(3):329-40. http://www.ncbi.nlm.nih.gov/pubmed/10337104
I. neotomae: (Also ; or now known as I spinaplpis – see Norris et al, 1997):
Keirans JE, Brown RN and Lane RS (1996) Ixodes (Ixodes) jellisoni and I. (I.) neotomae (Acari:Ixodidae): descriptions of the immature stages from California. J Med Entomol;33(3):319-27. http://www.ncbi.nlm.nih.gov/pubmed/8667376
Norris DE, Klompen JS, Keirans JE, Lane RS, Piesman J and Black WC 4th(1997) Taxonomic status of Ixodes neotomae and I. spinipalpis (Acari: Ixodidae) based on mitochondrial DNA evidence. J Med Entomol; 34(6):696-703. http://www.ncbi.nlm.nih.gov/pubmed/9439125
I. spinipalpis:
Dolan MC, Maupin GO, Panella NA, Golde WT, Piesman J (1997) Vector competence of Ixodes scapularis, I. spinipalpis, and Dermacentor andersoni (Acari:Ixodidae) in transmitting Borrelia burgdorferi, the etiologic agent of Lyme disease. J Med Entomol; 34(2):128-35. http://www.ncbi.nlm.nih.gov/pubmed/9103755
I. angustus:
Peavey CA, Lane RS and Damrow T (2000) Vector competence of Ixodes angustus (Acari: Ixodidae) for Borrelia burgdorferi sensu stricto. Exp Appl Acarol;24(1):77-84. http://www.ncbi.nlm.nih.gov/pubmed/10823359
I. minor:
(a) Rudenko N, Golovchenko M, Lin T, Gao L, Grubhoffer L and Oliver JH Jr (2009) Delineation of a new species of the Borrelia burgdorferi Sensu Lato Complex, Borrelia americana sp. nov. J Clin Microbiol;47(12):3875-80. Epub 2009 Oct 21. http://www.ncbi.nlm.nih.gov/pubmed/19846628
(b) Clark KL, Oliver JH Jr, Grego JM, James AM, Durden LA and Banks CW (2001) Host associations of ticks parasitizing rodents at Borrelia burgdorferi enzootic sites in South Carolina. J Parasitol;87(6):1379-86. http://www.ncbi.nlm.nih.gov/pubmed/11780825
I. muris:
Dolan MC, Lacombe EH and Piesman J (2000) Vector competence of Ixodes muris (Acari: Ixodidae) for Borrelia burgdorferi. J Med Entomol; 37(5):766-8. http://www.ncbi.nlm.nih.gov/pubmed/11004792
A. Americanum:
Schulze TL, Jordan RA, Schulze CJ, Mixon T and Papero M (2005) Relative encounter frequencies and prevalence of selected Borrelia, Ehrlichia, and Anaplasma infections in Amblyomma americanum and Ixodes scapularis (Acari: Ixodidae) ticks from central New Jersey. J Med Entomol;42(3):450-6. http://www.ncbi.nlm.nih.gov/pubmed/15962799
H. leporispalustris:
(a) Lane RS and Burgdorfer W (1988) Spirochetes in mammals and ticks (Acari: Ixodidae) from a focus of Lyme borreliosis in California. J Wildl Dis; 24(1):1-9. http://www.ncbi.nlm.nih.gov/pubmed/3280837
(b) Banerjee SN, Banerjee M, Fernando K, Dong MY, Smith JA and Cook D (1995) Isolation of Borrelia burgdorferi, the lyme disease spirochete, from rabbit ticks, haemaphysalis leporispalustris – Alberta. Can Commun Dis Rep;21(10):86-8.
http://www.ncbi.nlm.nih.gov/pubmed/7620455 Full copy at: http://www.jwildlifedis.org/cgi/reprint/24/1/1
(c) Nicholls TH.and Callister SM (1996) Lyme Disease (Borelia burgdorferi) Spirochetes in Ticks collected from birds in midwestern United states. Abstract from chapter in Journal of Medical Entomology; 33(3): 379-384. http://www.treesearch.fs.fed.us/pubs/11268
I. scapularis in Canada:
CanadaCommunicable Disease Report (1995) Vol 21-10. ISSN 1188-4169
http://www.collectionscanada.gc.ca/webarchives/20071127051546/http://www.phac-aspc.gc.ca/publicat/ccdr-rmtc/95pdf/cdr2110e.pdf
I. auritulus:
(a) Scott JD, Anderson JF and Durden LA (2011) Widespread dispersal of Borrelia burgdorferi-infected ticks collected from songbirds across Canada. J Parasitol Aug 24. [Epub ahead of print] http://www.ncbi.nlm.nih.gov/pubmed/21864130
(b) Scott JD, Lee MK, Fernando K, Durden LA, Jorgensen DR, Mak S and Morshed MG (2010) Detection of Lyme disease spirochete, Borrelia burgdorferi sensu lato, including three novel genotypes in ticks (Acari: Ixodidae) collected from songbirds (Passeriformes) across Canada. J Vector Ecol;35(1):124-39. http://www.ncbi.nlm.nih.gov/pubmed/20618658
(c) Morshed MG, Scott JD, Fernando K, Beati L, Mazerolle DF, Geddes G and Durden LA (2005) Migratory songbirds disperse ticks across Canada, and first isolation of the Lyme disease spirochete, Borrelia burgdorferi, from the avian tick, Ixodes auritulus. J Parasitol;91(4):780-90. http://www.ncbi.nlm.nih.gov/pubmed/17089744
I. ricinus: Well known vector
I. hexagonus:
(a) Toutoungi LN and Gern L (1993) Ability of transovarially and subsequent transstadially infected Ixodes hexagonus ticks to maintain and transmit Borrelia burgdorferi in the laboratory. Exp Appl Acarol;17(8):581-6. http://www.ncbi.nlm.nih.gov/pubmed/7628234
(b) Gern L, Rouvinez E, Toutoungi LN and Godfroid E (1997) Transmission cycles of Borrelia burgdorferi sensu lato involving Ixodes ricinus and/or I. hexagonus ticks and the European hedgehog, Erinaceus europaeus, in suburban and urban areas in Switzerland. Folia Parasitol (Praha) ;44(4):309-14. http://www.ncbi.nlm.nih.gov/pubmed/9437846
(c) Estrada-Pena A, Oteo JA, Estrada-Pena R, Gortazar C, Osacar JJ, Moreno JA and Castella J (1995) Borrelia burgdorferi sensu lato in ticks (Acari: Ixodidae) from two different foci in Spain. Exp Appl Acarol; 19(3):173-80. http://www.ncbi.nlm.nih.gov/pubmed/7634972
I. canisuga and I. frontalis:
Estrada-Pena A, Oteo JA, Estrada-Pena R, Gortazar C, Osacar JJ, Moreno JA and Castella J (1995) Borrelia burgdorferi sensu lato in ticks (Acari: Ixodidae) from two different foci in Spain. Exp Appl Acarol;19(3):173-80. http://www.ncbi.nlm.nih.gov/pubmed/7634972
I. trianguliceps:
(a) Gorelova NB, Korenberg EI, Kovalevskii JuV, Postic D and Baranton G (1996) The isolation of Borrelia from the tick Ixodes trianguliceps (Ixodidae) and the possible significance of this species in the epizootiology of ixodid tick-borne borrelioses. Parazitologiia;30(1):13-8. http://www.ncbi.nlm.nih.gov/pubmed/8975209
(b) Grigor’eva LA and Tret’kiakov KA (1998) Features of the parasitic system of Ixodid ticks--Borrelia--small mammals in the Russian Northwest. Parazitologiia;32(5):422-30. http://www.ncbi.nlm.nih.gov/pubmed/9859127
(c) Hubbard MJ, Baker AS and Cann KJ (1998) Distribution of Borrelia burgdorferi s.l. spirochaete DNA in British ticks (Argasidae and Ixodidae) since the 19th century, assessed by PCR. Med Vet Entomol;12(1):89-97. http://www.ncbi.nlm.nih.gov/pubmed/9513944
I. persulcatus: Well known vector
I. sinensis:
(a) Sun, Y, Xu, R and Cao, W (2003). Ixodes sinensis: competence as a vector to transmit the Lyme disease spirochete Borrelia garinii. Vector Borne Zoonotic Dis; 3(1), 39–44 http://www.ncbi.nlm.nih.gov/pubmed/12804379
(b) Sun Yi, Xu R, Ge EF and Cao W (2009) Natural infection of Borrelia afzelii in Ixodes sinensis and its parasitism hosts in forest areas of Huangshan, Anhui Province. Acta Parasitoogica et Medica Entomologica Sinica; 16 (3), 141-146. ISSN 1005-0507. http://www.cabdirect.org/abstracts/20103039526.html;jsessionid=A15D6311D71E4ECD6782B133C5A84006
I. ovatus:
Kawabata H, Masuzawa T Yanagihara Y (1993) Genomic analysis of Borrelia japonica sp. nov. isolated from Ixodes ovatus in Japan. Microbiol. Immunol;37(11) 843-848 http://www.ncbi.nlm.nih.gov/pubmed/7905183
I nipponensis: (Indirect reference)
Masuzawa T (2004) Terrestrial Distribution of the Lyme Borreliosis Agent Borrelia burgdorferi Sensu Lato in East Asia. Jpn J Infect Dis, 57(6); 229-235. http://www.ncbi.nlm.nih.gov/pubmed/15623946
I granulatus and H bispinosa:
(a) Wan K, Zhang Z, and Dou G (1998) Investigation on primary vectors of Borrelia burgdorferi in China. Chin J Epidemiol 19, 263–266. http://www.ncbi.nlm.nih.gov/pubmed/10322682
(b) Chao LL , Wu WJ, and Shih CM (2009) First Detection and Molecular Identification of Borrelia burgdorferi -like Spirochetes in Ixodes granulatus Ticks Collected on Kinmen Island of Taiwan. Am. J. Trop. Med. Hyg; 80(3): 389–394. http://www.ajtmh.org/content/80/3/389.full.pdf
(c) Hao Q, Hou X, Geng Z and Wan K (2011) Distribution of Borrelia burgdorferi Sensu Lato in China. J Clin Microbiol; 49(2): 647-650. http://www.ncbi.nlm.nih.gov/pubmed/21106783
H. flava:
(a) Ishiguro F, Takada N, Masuzawa T and Fukui T (2000) Prevalence of Lyme disease Borrelia spp. in ticks from migratory birds on the Japanese mainland. Appl Environ Microbiol;66(3):982-6. http://www.ncbi.nlm.nih.gov/pubmed/10698761
(b) Ishiquro F, Takada N and Masuzawa T (2005) Molecular evidence of the dispersal of Lyme disease Borrelia from the Asian Continent to Japan via migratory birds. Jpn J Infect Dis; 58(3):184-6. http://www.ncbi.nlm.nih.gov/pubmed/15973014
H. longicornis
(a) Chu CY, Jiang BG, Liu W, Zhao QM, Wu XM, Zhang PH, Zhan H and Cao WC (2008). Presence of pathogenic Borrelia burgdorferi sensu lato in ticks and rodents in Zhejiang, south-east China. J Med Microbiol;57( 8):980-5 http://www.ncbi.nlm.nih.gov/pubmed/18628499
(b) Chu CY, Liu W, Jiang BG, Wang DM, Jiang WJ, Zhao QM, Zhang PH, Wang ZX, Tang GP, Yang H and Cao WC (2008) Novel Genospecies of Borrelia burgdorferi Sensu Lato from Rodents and Ticks in Southwestern China. J Clin Microbiol; 46(9):3130-3 http://www.ncbi.nlm.nih.gov/pubmed/18614645
(c) Sun J, Liu Q, Lu L, Ding G, Guo J, Fu G, Zhang J, Meng F, Wu H, Song X, Ren D, Li D, Guo Y, Wang J, Li G, Liu J and Lin H (2008) Coinfection with four genera of bacteria (Borrelia, Bartonella, Anaplasma, and Ehrlichia) in Haemaphysalis longicornis and Ixodes sinensis ticks from China. Vector Borne Zoonotic Dis; 8(6): 791-5. http://www.ncbi.nlm.nih.gov/pubmed/18637722
(d) Meng Z, Jiang LP, Lu QY, Cheng SY, Ye JL and Zhan L (2008) Detection of co-infection with Lyme spirochetes and spotted fever group rickettsiae in a group of Haemaphysalis longicornis. Zhonghua Liu Xing Bing Xue Za Zhi; 29 (12): 1217–1220. http://www.ncbi.nlm.nih.gov/pubmed/19173967
I. columnae, I. tanuki, I. turdis; (Indirect reference):
Masuzawa T (2004) Terrestrial Distribution of the Lyme Borreliosis Agent Borrelia burgdorferi Sensu Lato in East Asia. Jpn J Infect Dis, 57(6); 229-235. http://www.ncbi.nlm.nih.gov/pubmed/15623946
I. tanuki, I. turdis: (Indirect reference)
Saito K, Ito T, Asashima N, Ohno M, Nagai R, Fujita H, Koizumi N, Takano A, Watanabe H and Kawabata H (2007) Case Report: Borrelia valaisiana Infection in a Japanese Man Associated with Traveling to Foreign Countries. Am J Trop Med Hyg; 77(6): 1124–1127. http://www.ajtmh.org/content/77/6/1124.full.pdf
I Uriae:
(a) Olsen B, Jaenson TG, Noppa L, Bunikis J, Bergstrom S (1993) A Lyme borreliosis cycle in seabirds and Ixodes uriae ticks. Nature; 362:340-342. http://www.ncbi.nlm.nih.gov/pubmed/8455718
(b) Olsen B, Duffy DC, Jaenson TG, Gylfe A, Bonnedahl J and Bergstrom S (1995) Transhemispheric exchange of Lyme disease spirochetes by seabirds. J Clin Microbiol; 33:3270-3274 http://www.ncbi.nlm.nih.gov/pubmed/8586715
Tick/Vector References Further to the above:
(1) Maupin GO, Gage KL, Piesman J, Montenieri J, Sviat SL, VanderZanden L, Happ CM, Dolan M and Johnson BJ (1994) Discovery of an enzootic cycle of Borrelia burgdorferi in Neotoma mexicana and Ixodes spinipalpis from northern Colorado, an area where Lyme disease is nonendemic. J Infect Dis; 170(3):636-43. http://www.ncbi.nlm.nih.gov/pubmed/8077722
(2) Postic D, Ras NM, Lane RS, Hendson M and Baranton G (1998) Expanded diversity among Californian borrelia isolates and description of Borrelia bissettii sp. nov. (formerly Borrelia group DN127). J Clin Microbiol; 36(12):3497-504. http://www.ncbi.nlm.nih.gov/pubmed/9817861
(3) Steere AC, Coburn J and Glickstein L (2004) The emergence of Lyme disease. J Clin Invest; 113(8):1093-101.
http://www.ncbi.nlm.nih.gov/pubmed/15085185
(4) Burkot TR, Maupin Go, Schneider BS, Denatale C, Happ CM, Rutherford JS and Zeidner NS (2001) Use of a sentinel host
system to study the questing behavior of Ixodes spinipalpis and its role in the transmission of Borrelia bissettii, human granulocytic ehrlichiosis, and Babesia microti. Am J Trop Med Hyg;65(4):293-9. http://www.ncbi.nlm.nih.gov/pubmed/11693872
How Lyme is Transmitted and Maintained in the Environment. Ticks and Human Disease
(30) Gern L and Humair PF (2002) Ecology of Borrelia burgdorferi senso lato in Europe. Lyme Borreliosis : Biology, Epidemiology and Control 6: 149-174 http://doc.rero.ch/lm.php?url=1000,43,4,20060515171152-TL/1_Gern_Lise_-_Ecology_of_Borrelia_burgdorferi_20060515.pdf
(31) Kurtenbach K, De Michelis S, Sewell HS, Etti S, Schafer SM, Hails R, Collares-Pereira M, Santos-Reis M, Hanincova K, Labuda M, Bormane A and Donaghy M (2001) Distinct Combinations of Borrelia burgdorferi Sensu Lato Genospecies Found in Individual Questing Ticks from Europe. Appl Environ Microbiol; 67(10):4926-9. http://www.ncbi.nlm.nih.gov/pubmed/11571205
(32) Vector-Borne Diseases ; e-notes: http://www.enotes.com/vector-borne-diseases-reference/vector-borne-diseases
(33) Depart Entomology, Uni of California: http://entomology.ucdavis.edu/faculty/rbkimsey/tickbio.html
Lyme Disease Tick Vectors
I. scapularis and I. pacificus: Well known vectors
I. dentatus:
(a) Anderson JF, Magnarelli LA and Stafford KC 3rd (1990) Bird-feeding ticks transstadially transmit Borrelia burgdorferi that infect Syrian hamsters. J Wildl Dis; 26 (1):1-10. http://www.ncbi.nlm.nih.gov/pubmed/2304189
(b) Indirect ref: Masuzawa T (2004) Terrestrial Distribution of the Lyme Borreliosis Agent Borrelia burgdorferi Sensu Lato in East Asia. Jpn J Infect Dis, 57(6); 229-235. http://www.ncbi.nlm.nih.gov/pubmed/15623946
I. affinis:
Maggi RG, Reichelt S, Toliver M and Engber B (2010) Borrelia species in Ixodes affinis and Ixodes scapularis ticks collected from the coastal plain of North Carolina. Ticks Tick Borne Dis;1(4):168-71. Epub 2010 Oct 20. http://www.ncbi.nlm.nih.gov/pubmed/21771524
I. jellisoni:
Lane RS, Peavey CA, Padgett KA and Hendson M (1999) Life history of Ixodes (Ixodes) jellisoni (Acari: Ixodidae) and its vector competence for Borrelia burgdorferi sensu lato. J Med Entomol;36(3):329-40. http://www.ncbi.nlm.nih.gov/pubmed/10337104
I. neotomae: (Also ; or now known as I spinaplpis – see Norris et al, 1997):
Keirans JE, Brown RN and Lane RS (1996) Ixodes (Ixodes) jellisoni and I. (I.) neotomae (Acari:Ixodidae): descriptions of the immature stages from California. J Med Entomol;33(3):319-27. http://www.ncbi.nlm.nih.gov/pubmed/8667376
Norris DE, Klompen JS, Keirans JE, Lane RS, Piesman J and Black WC 4th(1997) Taxonomic status of Ixodes neotomae and I. spinipalpis (Acari: Ixodidae) based on mitochondrial DNA evidence. J Med Entomol; 34(6):696-703. http://www.ncbi.nlm.nih.gov/pubmed/9439125
I. spinipalpis:
Dolan MC, Maupin GO, Panella NA, Golde WT, Piesman J (1997) Vector competence of Ixodes scapularis, I. spinipalpis, and Dermacentor andersoni (Acari:Ixodidae) in transmitting Borrelia burgdorferi, the etiologic agent of Lyme disease. J Med Entomol; 34(2):128-35. http://www.ncbi.nlm.nih.gov/pubmed/9103755
I. angustus:
Peavey CA, Lane RS and Damrow T (2000) Vector competence of Ixodes angustus (Acari: Ixodidae) for Borrelia burgdorferi sensu stricto. Exp Appl Acarol;24(1):77-84. http://www.ncbi.nlm.nih.gov/pubmed/10823359
I. minor:
(a) Rudenko N, Golovchenko M, Lin T, Gao L, Grubhoffer L and Oliver JH Jr (2009) Delineation of a new species of the Borrelia burgdorferi Sensu Lato Complex, Borrelia americana sp. nov. J Clin Microbiol;47(12):3875-80. Epub 2009 Oct 21. http://www.ncbi.nlm.nih.gov/pubmed/19846628
(b) Clark KL, Oliver JH Jr, Grego JM, James AM, Durden LA and Banks CW (2001) Host associations of ticks parasitizing rodents at Borrelia burgdorferi enzootic sites in South Carolina. J Parasitol;87(6):1379-86. http://www.ncbi.nlm.nih.gov/pubmed/11780825
I. muris:
Dolan MC, Lacombe EH and Piesman J (2000) Vector competence of Ixodes muris (Acari: Ixodidae) for Borrelia burgdorferi. J Med Entomol; 37(5):766-8. http://www.ncbi.nlm.nih.gov/pubmed/11004792
A. Americanum:
Schulze TL, Jordan RA, Schulze CJ, Mixon T and Papero M (2005) Relative encounter frequencies and prevalence of selected Borrelia, Ehrlichia, and Anaplasma infections in Amblyomma americanum and Ixodes scapularis (Acari: Ixodidae) ticks from central New Jersey. J Med Entomol;42(3):450-6. http://www.ncbi.nlm.nih.gov/pubmed/15962799
H. leporispalustris:
(a) Lane RS and Burgdorfer W (1988) Spirochetes in mammals and ticks (Acari: Ixodidae) from a focus of Lyme borreliosis in California. J Wildl Dis; 24(1):1-9. http://www.ncbi.nlm.nih.gov/pubmed/3280837
(b) Banerjee SN, Banerjee M, Fernando K, Dong MY, Smith JA and Cook D (1995) Isolation of Borrelia burgdorferi, the lyme disease spirochete, from rabbit ticks, haemaphysalis leporispalustris – Alberta. Can Commun Dis Rep;21(10):86-8.
http://www.ncbi.nlm.nih.gov/pubmed/7620455 Full copy at: http://www.jwildlifedis.org/cgi/reprint/24/1/1
(c) Nicholls TH.and Callister SM (1996) Lyme Disease (Borelia burgdorferi) Spirochetes in Ticks collected from birds in midwestern United states. Abstract from chapter in Journal of Medical Entomology; 33(3): 379-384. http://www.treesearch.fs.fed.us/pubs/11268
I. scapularis in Canada:
CanadaCommunicable Disease Report (1995) Vol 21-10. ISSN 1188-4169
http://www.collectionscanada.gc.ca/webarchives/20071127051546/http://www.phac-aspc.gc.ca/publicat/ccdr-rmtc/95pdf/cdr2110e.pdf
I. auritulus:
(a) Scott JD, Anderson JF and Durden LA (2011) Widespread dispersal of Borrelia burgdorferi-infected ticks collected from songbirds across Canada. J Parasitol Aug 24. [Epub ahead of print] http://www.ncbi.nlm.nih.gov/pubmed/21864130
(b) Scott JD, Lee MK, Fernando K, Durden LA, Jorgensen DR, Mak S and Morshed MG (2010) Detection of Lyme disease spirochete, Borrelia burgdorferi sensu lato, including three novel genotypes in ticks (Acari: Ixodidae) collected from songbirds (Passeriformes) across Canada. J Vector Ecol;35(1):124-39. http://www.ncbi.nlm.nih.gov/pubmed/20618658
(c) Morshed MG, Scott JD, Fernando K, Beati L, Mazerolle DF, Geddes G and Durden LA (2005) Migratory songbirds disperse ticks across Canada, and first isolation of the Lyme disease spirochete, Borrelia burgdorferi, from the avian tick, Ixodes auritulus. J Parasitol;91(4):780-90. http://www.ncbi.nlm.nih.gov/pubmed/17089744
I. ricinus: Well known vector
I. hexagonus:
(a) Toutoungi LN and Gern L (1993) Ability of transovarially and subsequent transstadially infected Ixodes hexagonus ticks to maintain and transmit Borrelia burgdorferi in the laboratory. Exp Appl Acarol;17(8):581-6. http://www.ncbi.nlm.nih.gov/pubmed/7628234
(b) Gern L, Rouvinez E, Toutoungi LN and Godfroid E (1997) Transmission cycles of Borrelia burgdorferi sensu lato involving Ixodes ricinus and/or I. hexagonus ticks and the European hedgehog, Erinaceus europaeus, in suburban and urban areas in Switzerland. Folia Parasitol (Praha) ;44(4):309-14. http://www.ncbi.nlm.nih.gov/pubmed/9437846
(c) Estrada-Pena A, Oteo JA, Estrada-Pena R, Gortazar C, Osacar JJ, Moreno JA and Castella J (1995) Borrelia burgdorferi sensu lato in ticks (Acari: Ixodidae) from two different foci in Spain. Exp Appl Acarol; 19(3):173-80. http://www.ncbi.nlm.nih.gov/pubmed/7634972
I. canisuga and I. frontalis:
Estrada-Pena A, Oteo JA, Estrada-Pena R, Gortazar C, Osacar JJ, Moreno JA and Castella J (1995) Borrelia burgdorferi sensu lato in ticks (Acari: Ixodidae) from two different foci in Spain. Exp Appl Acarol;19(3):173-80. http://www.ncbi.nlm.nih.gov/pubmed/7634972
I. trianguliceps:
(a) Gorelova NB, Korenberg EI, Kovalevskii JuV, Postic D and Baranton G (1996) The isolation of Borrelia from the tick Ixodes trianguliceps (Ixodidae) and the possible significance of this species in the epizootiology of ixodid tick-borne borrelioses. Parazitologiia;30(1):13-8. http://www.ncbi.nlm.nih.gov/pubmed/8975209
(b) Grigor’eva LA and Tret’kiakov KA (1998) Features of the parasitic system of Ixodid ticks--Borrelia--small mammals in the Russian Northwest. Parazitologiia;32(5):422-30. http://www.ncbi.nlm.nih.gov/pubmed/9859127
(c) Hubbard MJ, Baker AS and Cann KJ (1998) Distribution of Borrelia burgdorferi s.l. spirochaete DNA in British ticks (Argasidae and Ixodidae) since the 19th century, assessed by PCR. Med Vet Entomol;12(1):89-97. http://www.ncbi.nlm.nih.gov/pubmed/9513944
I. persulcatus: Well known vector
I. sinensis:
(a) Sun, Y, Xu, R and Cao, W (2003). Ixodes sinensis: competence as a vector to transmit the Lyme disease spirochete Borrelia garinii. Vector Borne Zoonotic Dis; 3(1), 39–44 http://www.ncbi.nlm.nih.gov/pubmed/12804379
(b) Sun Yi, Xu R, Ge EF and Cao W (2009) Natural infection of Borrelia afzelii in Ixodes sinensis and its parasitism hosts in forest areas of Huangshan, Anhui Province. Acta Parasitoogica et Medica Entomologica Sinica; 16 (3), 141-146. ISSN 1005-0507. http://www.cabdirect.org/abstracts/20103039526.html;jsessionid=A15D6311D71E4ECD6782B133C5A84006
I. ovatus:
Kawabata H, Masuzawa T Yanagihara Y (1993) Genomic analysis of Borrelia japonica sp. nov. isolated from Ixodes ovatus in Japan. Microbiol. Immunol;37(11) 843-848 http://www.ncbi.nlm.nih.gov/pubmed/7905183
I nipponensis: (Indirect reference)
Masuzawa T (2004) Terrestrial Distribution of the Lyme Borreliosis Agent Borrelia burgdorferi Sensu Lato in East Asia. Jpn J Infect Dis, 57(6); 229-235. http://www.ncbi.nlm.nih.gov/pubmed/15623946
I granulatus and H bispinosa:
(a) Wan K, Zhang Z, and Dou G (1998) Investigation on primary vectors of Borrelia burgdorferi in China. Chin J Epidemiol 19, 263–266. http://www.ncbi.nlm.nih.gov/pubmed/10322682
(b) Chao LL , Wu WJ, and Shih CM (2009) First Detection and Molecular Identification of Borrelia burgdorferi -like Spirochetes in Ixodes granulatus Ticks Collected on Kinmen Island of Taiwan. Am. J. Trop. Med. Hyg; 80(3): 389–394. http://www.ajtmh.org/content/80/3/389.full.pdf
(c) Hao Q, Hou X, Geng Z and Wan K (2011) Distribution of Borrelia burgdorferi Sensu Lato in China. J Clin Microbiol; 49(2): 647-650. http://www.ncbi.nlm.nih.gov/pubmed/21106783
H. flava:
(a) Ishiguro F, Takada N, Masuzawa T and Fukui T (2000) Prevalence of Lyme disease Borrelia spp. in ticks from migratory birds on the Japanese mainland. Appl Environ Microbiol;66(3):982-6. http://www.ncbi.nlm.nih.gov/pubmed/10698761
(b) Ishiquro F, Takada N and Masuzawa T (2005) Molecular evidence of the dispersal of Lyme disease Borrelia from the Asian Continent to Japan via migratory birds. Jpn J Infect Dis; 58(3):184-6. http://www.ncbi.nlm.nih.gov/pubmed/15973014
H. longicornis
(a) Chu CY, Jiang BG, Liu W, Zhao QM, Wu XM, Zhang PH, Zhan H and Cao WC (2008). Presence of pathogenic Borrelia burgdorferi sensu lato in ticks and rodents in Zhejiang, south-east China. J Med Microbiol;57( 8):980-5 http://www.ncbi.nlm.nih.gov/pubmed/18628499
(b) Chu CY, Liu W, Jiang BG, Wang DM, Jiang WJ, Zhao QM, Zhang PH, Wang ZX, Tang GP, Yang H and Cao WC (2008) Novel Genospecies of Borrelia burgdorferi Sensu Lato from Rodents and Ticks in Southwestern China. J Clin Microbiol; 46(9):3130-3 http://www.ncbi.nlm.nih.gov/pubmed/18614645
(c) Sun J, Liu Q, Lu L, Ding G, Guo J, Fu G, Zhang J, Meng F, Wu H, Song X, Ren D, Li D, Guo Y, Wang J, Li G, Liu J and Lin H (2008) Coinfection with four genera of bacteria (Borrelia, Bartonella, Anaplasma, and Ehrlichia) in Haemaphysalis longicornis and Ixodes sinensis ticks from China. Vector Borne Zoonotic Dis; 8(6): 791-5. http://www.ncbi.nlm.nih.gov/pubmed/18637722
(d) Meng Z, Jiang LP, Lu QY, Cheng SY, Ye JL and Zhan L (2008) Detection of co-infection with Lyme spirochetes and spotted fever group rickettsiae in a group of Haemaphysalis longicornis. Zhonghua Liu Xing Bing Xue Za Zhi; 29 (12): 1217–1220. http://www.ncbi.nlm.nih.gov/pubmed/19173967
I. columnae, I. tanuki, I. turdis; (Indirect reference):
Masuzawa T (2004) Terrestrial Distribution of the Lyme Borreliosis Agent Borrelia burgdorferi Sensu Lato in East Asia. Jpn J Infect Dis, 57(6); 229-235. http://www.ncbi.nlm.nih.gov/pubmed/15623946
I. tanuki, I. turdis: (Indirect reference)
Saito K, Ito T, Asashima N, Ohno M, Nagai R, Fujita H, Koizumi N, Takano A, Watanabe H and Kawabata H (2007) Case Report: Borrelia valaisiana Infection in a Japanese Man Associated with Traveling to Foreign Countries. Am J Trop Med Hyg; 77(6): 1124–1127. http://www.ajtmh.org/content/77/6/1124.full.pdf
I Uriae:
(a) Olsen B, Jaenson TG, Noppa L, Bunikis J, Bergstrom S (1993) A Lyme borreliosis cycle in seabirds and Ixodes uriae ticks. Nature; 362:340-342. http://www.ncbi.nlm.nih.gov/pubmed/8455718
(b) Olsen B, Duffy DC, Jaenson TG, Gylfe A, Bonnedahl J and Bergstrom S (1995) Transhemispheric exchange of Lyme disease spirochetes by seabirds. J Clin Microbiol; 33:3270-3274 http://www.ncbi.nlm.nih.gov/pubmed/8586715
Tick/Vector References Further to the above:
(1) Maupin GO, Gage KL, Piesman J, Montenieri J, Sviat SL, VanderZanden L, Happ CM, Dolan M and Johnson BJ (1994) Discovery of an enzootic cycle of Borrelia burgdorferi in Neotoma mexicana and Ixodes spinipalpis from northern Colorado, an area where Lyme disease is nonendemic. J Infect Dis; 170(3):636-43. http://www.ncbi.nlm.nih.gov/pubmed/8077722
(2) Postic D, Ras NM, Lane RS, Hendson M and Baranton G (1998) Expanded diversity among Californian borrelia isolates and description of Borrelia bissettii sp. nov. (formerly Borrelia group DN127). J Clin Microbiol; 36(12):3497-504. http://www.ncbi.nlm.nih.gov/pubmed/9817861
(3) Steere AC, Coburn J and Glickstein L (2004) The emergence of Lyme disease. J Clin Invest; 113(8):1093-101.
http://www.ncbi.nlm.nih.gov/pubmed/15085185
(4) Burkot TR, Maupin Go, Schneider BS, Denatale C, Happ CM, Rutherford JS and Zeidner NS (2001) Use of a sentinel host
system to study the questing behavior of Ixodes spinipalpis and its role in the transmission of Borrelia bissettii, human granulocytic ehrlichiosis, and Babesia microti. Am J Trop Med Hyg;65(4):293-9. http://www.ncbi.nlm.nih.gov/pubmed/11693872
